
Javier Ortiz-Tudela y Juan Lupiáñez
Centro de Investigación Mente, Cerebro y Comportamiento, Universidad de Granada, España

(dp) Horatio Henry Couldery, «The unexpected guest», 1874.
Si pensamos en la función cerebral como fundamentalmente predictora del ambiente, nuestro sistema cognitivo debería contar con herramientas que le permitieran 1) establecer esas predicciones y reaccionar de acuerdo con ellas, y 2) detectar cuándo esas predicciones no se cumplen para disparar un mecanismo que lleve a adquirir nueva información para minimizar el error en las futuras predicciones. No obstante, aislar en el laboratorio estos dos tipos de procesos es bastante complicado. El estudio que presentamos aquí muestra una manera de separar la respuesta de detección (saber que algo está presente) de la de identificación (saber qué es ese algo). La primera serviría como señal de alerta ante la falta de predicción y la segunda sería un reflejo de esas predicciones hechas por el cerebro.
La concepción del cerebro como un órgano que acumula experiencias y cuya principal función es la de predecir y anticipar las situaciones a las que el organismo se enfrenta ha sido una aproximación común a distintas ramas de la investigación en psicología: desde los modelos clásicos de aprendizaje (Rescorla y Wagner, 1972) hasta modelos más actuales de funcionamiento neuronal (Bastos y col., 2012). Según esta aproximación nuestro cerebro utilizaría los conocimientos almacenados en memoria y la información presente para crear una representación integrada del mundo. A partir de esta representación sería posible anticipar (probabilísticamente) los acontecimientos futuros y así responder de manera rápida y precisa cuando estas predicciones se cumplen. Esta preparación supondría, por contra, una peor respuesta cuando el ambiente se comporta de manera imprevista. Bajo esta perspectiva se pueden explicar numerosos fenómenos de la psicología experimental clásica, tales como el “priming” (el mejor procesamiento de un estímulo si éste ha sido precedido por otro relacionado con él) o la interpretación de estímulos degradados (donde un buen ajuste entre los distintos elementos presentes en una escena facilita su identificación).
No obstante, un sistema que contara sólo con mecanismos encargados de predecir y reaccionar de acuerdo con esas predicciones se encontraría indefenso ante situaciones en las que se enfrentara con información inconsistente. Un mundo probabilístico como es el nuestro fuerza a un sistema que pretenda adaptarse a él a dotarse de herramientas que le permitan reaccionar eficientemente ante situaciones novedosas. Es, por tanto, inevitable pensar que nuestro cerebro cuente con algún instrumento para detectar esas incongruencias, o falta de coincidencia (bien entre los elementos presentes perceptivamente, o entre éstos y las predicciones hechas sobre ellos), y que desencadene los procesos necesarios para reaccionar ante ellas de manera adecuada.
Un posible candidato para esta tarea sería un componente atencional que ha sido relacionado frecuentemente con la actividad en regiones anteriores de la corteza cingulada y que dispararía una alerta cuando las estrategias automatizadas son ineficaces y se necesita una respuesta más específica y deliberada (Carter, 1998). Este procesamiento requeriría de la integración de la información presente en los aprendizajes previamente establecidos (actividad vinculada principalmente a regiones prefrontales; Miller y Cohen, 2001). La finalidad última de este proceso sería optimizar los pronósticos futuros, minimizando así el error de predicción y maximizando la eficiencia en la respuesta.
Si asumimos, por tanto, que nuestro sistema cognitivo cuenta con dos mecanismos, uno para detectar inconsistencias y otro para la integración de esa información novedosa, y que éstos interactúan y funcionan en paralelo, pero que tienen características y temporalidad distintas, deberíamos ser capaces de disociarlos variando las exigencias que hagamos al sistema. En esta línea, en un estudio reciente de nuestro laboratorio (LaPointe, Lupiáñez, y Milliken, 2013) se pretendía separar estos dos procesos en una misma tarea. LaPointe y col. (2013) se valieron de la tarea de detección de cambio, en la que manipularon la concordancia entre los objetos a detectar y el contexto de la imagen (véase la Figura 1 para un ejemplo de las imágenes usadas). En esta tarea se presentaba a los participantes un bucle de dos imágenes idénticas salvo por uno de los objetos. Entre las presentaciones de ambas imágenes se intercalaba una pantalla en blanco para impedir la captura atencional por parte del estímulo cambiante (Figura 2). Los participantes debían pulsar una tecla tan pronto como se dieran cuenta de que había un cambio (esto es, al detectar un objeto diferente de una imagen a la otra). Al hacer esto, la presentación finalizaba y tenían que identificar el objeto detectado.

Figura 1.- Ejemplo de imágenes utilizadas en el estudio de LaPointe y col. (2013). La saliencia de las imágenes fue manipulada de tal manera que las parejas de estímulos consistentes-inconsistentes no presentaran diferencia.
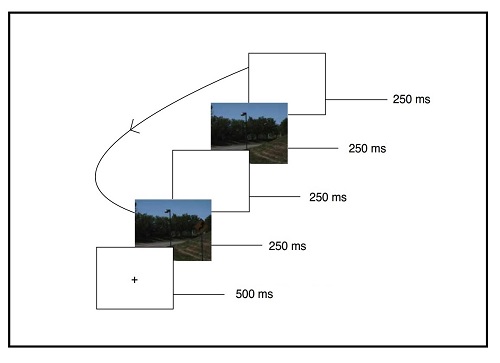
Figura 2.- Ejemplo de la sucesión de imágenes utilizado en la tarea de Detección de Cambio de LaPointe y col. (2013). Los participantes debían pulsar la barra espaciadora tan pronto como detectaran un cambio entre las imágenes, y a continuación debían identificar en qué consistía el cambio, es decir, cuál era el objeto que cambiaba de una imagen a otra. Al presentar la imagen en blanco entre una imagen y otra es muy difícil darse cuenta del cambio.
Los resultados fueron claros: aquellos objetos incongruentes con respecto a su contexto (p.ej., una señal de no fumar que aparece en el salón de una casa, como en la Figura 1) fueron detectados con mayor rapidez y en un mayor número de ocasiones que los congruentes (p.ej., esa misma señal en la terraza de un restaurante). De hecho, en el contexto congruente más del 10% de los objetos no fueron detectados en los 19 segundos que, como máximo, duraba la alternancia de imágenes. Sin embargo, esos mismos objetos, una vez detectados en el contexto congruente, se identificaron mejor que en el contexto incongruente (Figura 3). Pareciera como si el cerebro preactivara (en función de la experiencia previa) los conceptos relacionados con la escena. Esto haría más difícil detectarlos, porque sería como si siempre estuvieran presentes (cuando no están, los pondría el cerebro con su predictividad semántica), y facilitaría el reconocerlos cuando se detectan. Al contrario, aquellos objetos que no encajan con la escena serán más salientes, más llamativos (lo que ayudará a la hora de detectarlos como novedosos), pero más difíciles de identificar. Dado que la falta de encaje se produce a nivel semántico, estos resultados demuestran que hay procesamiento semántico de los objetos incongruentes antes de su identificación consciente.
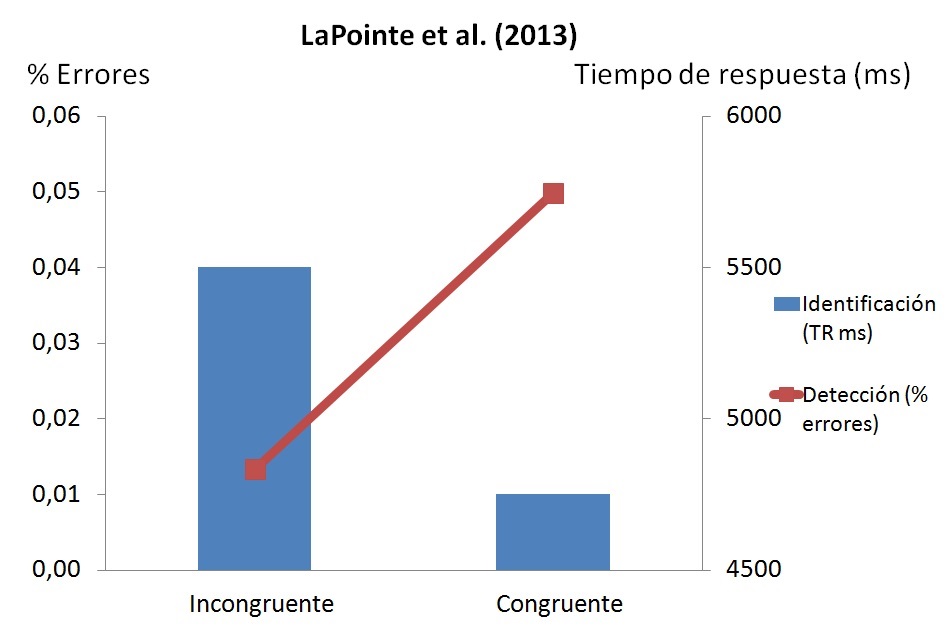
Figura 3.- Resultados del estudio de LaPointe y col. (2013): puntuaciones de identificación y detección en los dos contextos utilizados. Obsérvese que el tiempo empleado en detectar los cambios consistentes era mucho más elevado que el empleado en detectar los cambios inconsistentes, pero el número de errores cometidos en la identificación de los objetos detectados era menor en los objetos consistentes. Por tanto, esta congruencia semántica entre los objetos y su contexto perjudica la detección, pero facilita la identificación.
Esta investigación apoya la hipótesis de la existencia de dos procesos diferenciados: uno encargado de la integración de la información presente para propiciar un procesamiento fluido y otro encargado de detectar posibles incoherencias. El primero facilitaría un modo de actuación más automático (y, como consecuencia, eficiente), mientras que el segundo serviría para alertar de la necesidad de control. Esto reflejaría, por tanto, un funcionamiento dual del cerebro de acuerdo con dos modos de actuación: 1) un modo por defecto, de generación de predicciones, integración de información y preactivación de respuestas frecuentemente asociadas a esos estímulos (del que la conducta de identificación sería un ejemplo); y, 2) otro de respuestas rápidas e inespecíficas (conducta de detección), disparado frente a incongruencias y que, presumiblemente, conlleva un nuevo aprendizaje orientado a minimizar el error en las predicciones futuras.
Esta idea es compatible con las teorías clásicas de aprendizaje para las que la congruencia empeoraría el aprendizaje (no es necesario aprender de aquello que ya se sabe) y la incongruencia serviría como impulso para adquirir asociaciones nuevas (sólo se aprende de aquello novedoso o informativo). Esta aplicación de principios clásicos a la concepción del cerebro como un órgano predictor del ambiente representa un amplio marco conceptual a partir del cual es posible generar gran cantidad de nuevas investigaciones
Referencias
Bastos, A. M., Usrey, W. M., Adams, R. A., Mangun, G. R., Fries, P., y Friston, K. J. (2012). Canonical microcircuits for predictive coding. Neuron, 76(4), 695–711.
Carter, C. S. (1998). Anterior cingulate cortex, error detection, and the online monitoring of performance. Science, 280(5364), 747–749.
Clark, A. (2013). Whatever next? Predictive brains, situated agents, and the future of cognitive science. Behavioral and Brain Sciences, 36(3), 181–204.
LaPointe, M. R. P., Lupianez, J., y Milliken, B. (2013). Context congruency effects in change detection: Opposing effects on detection and identification. Visual Cognition, 21, 99–122.
Miller, E. K., y Cohen, J. D. (2001). An integrative theory of prefrontal cortex function. Annual Review of Neuroscience, 24, 167–202.
Rescorla, R. A., y Wagner, A. R. (1972). A theory of Pavlovian conditioning: Variations in the effectiveness of reinforcement and nonreinforcement. En: A. H. Black & W. F. Prokasy (Eds.), Classical Conditioning II: Current Research and Theory (pp. 64–99). New York, USA: Appleton-Century-Crofts.
Manuscrito recibido el 2 de marzo de 2015.
Aceptado el 16 de abril de 2015.
