
Francisco Aboitiz
Departamento de Psiquiatría, Pontificia Universidad Católica de Chile, Chile

NARA. "The Conversation". Dominio público.
En este artículo expongo dos interpretaciones recientes acerca del origen del lenguaje: la primera hace énfasis en el desarrollo de circuitos auditivo-vocales en los homínidos primitivos. La segunda apunta al desarrollo previo de un lenguaje de signos, el cual eventualmente dio lugar al lenguaje hablado. La superposición de los circuitos involucrados en ambas funciones indica que, desde muy temprano, las señales comunicativas fueron multimodales, incluyendo tanto vocalizaciones como gestos. El lenguaje hablado se desarrolló debido a la expansión de los circuitos auditivo-vocales, los cuales posiblemente invadieron elementos neuronales previamente involucrados en otras funciones, como los de control manual.
El lenguaje es una de las características definitorias de nuestra especie. ¿Cómo surgió en nuestro cerebro esta capacidad y qué procesos biológicos subyacen a ella? Estas son preguntas fundamentales acerca de la evolución humana, para las que aún no hay una respuesta definitiva. Sin embargo, han surgido recientemente algunas luces en este tema, gracias a nuevas técnicas tractográficas que permiten delinear los circuitos neuronales asociados al lenguaje (por ejemplo, Frey y cols., 2008). Éstos se localizan en el hemisferio izquierdo de la mayoría de las personas, e involucran regiones de la corteza auditiva en el lóbulo temporal (área de Wernicke), la región temporoparietal y la región prefrontal ventromedial (área de Broca), que se conecta con la región premotora adyacente que controla los movimientos del sistema vocal. Se han propuesto dos vías principales (dorsal y ventral) que conectan estas áreas (véase la Figura 1). La vía dorsal participa en la articulación vocal e involucra tractos que conectan la corteza auditiva posterior con la región de Broca, incluyendo el lóbulo parietal inferior. La vía ventral conecta la corteza auditiva anterior con la región de Broca anterior a través del lóbulo temporal anterior. Este tracto participa en la identificación de los sonidos, y ha sido asociado a procesos semánticos y léxicos.
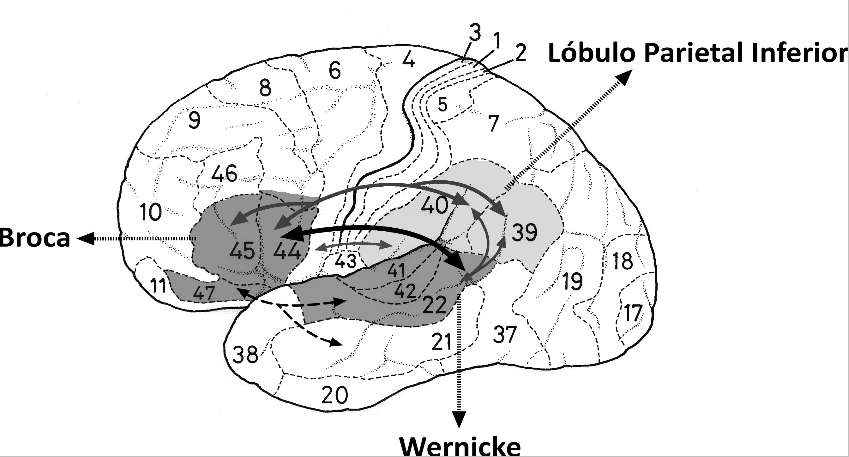
Figura 1.- Esta figura muestra la disposición de algunos tractos que conectan las regiones posteriores del lenguaje (área de Wernicke) con las regiones anteriores (área de Broca) en el hemisferio izquierdo humano. Las flechas en gris indican una proyección que pasa por el lóbulo parietal inferior; la flecha negra, mas oscura corresponde al fascículo arqueado, que se propone que conecta ambas regiones directamente; y las flechas segmentadas corresponden a la vía ventral que conecta las regiones auditivas anteriores con la región de Broca. (cc) F. Aboitiz.
En los primates no humanos, la comunicación vocal es rígida y estereotipada en su estructura, a diferencia de los humanos que tenemos un lenguaje casi ilimitado en sus expresiones. Esto es, en parte, gracias a nuestras capacidades de aprendizaje vocal. Un elemento de relevancia en el aprendizaje vocal es la memoria de trabajo verbal, que permite mantener activa la representación de secuencias fonológicas (p.ej., letras o números) por un breve tiempo. Los niños con más capacidad de memoria de trabajo fonológica eventualmente desarrollan vocabularios más amplios. Alan Baddeley (Baddeley y cols., 1988), quien acuñó el concepto de memoria de trabajo, llamaba “bucle fonológico” (phonological loop) al sistema sensoriomotor involucrado en este tipo de memoria. La memoria de trabajo fonológica se ha asociado predominante, pero no exclusivamente, a la activación de la vía dorsal mencionada arriba. Nuestro argumento original, en 1997, fue que en el cerebro del mono ya se observarían proyecciones desde el lóbulo parietal inferior y las áreas auditivas hacia la corteza prefrontal, que pueden ser consideradas como un rudimento de la vía dorsal asociada al bucle fonológico en humanos (Aboitiz y García, 1997). Como señalé arriba, esta hipótesis fue confirmada en estudios posteriores, con disponibilidad de nuevas tecnologías (entre otros, Frey y cols., 2008; Rilling y cols., 2008). Este circuito habría adquirido mayor prominencia en algún momento en la evolución de los homínidos, permitiendo el aprendizaje de vocalizaciones cada vez más complejas. La diferenciación de este circuito fonológico representó una etapa clave en la evolución de la humanidad, ya que permitió el desarrollo del habla, con todas las consecuencias cognitivas y sociales que esto pudo tener (Aboitiz, 2012).
Paradójicamente, en el mono las proyecciones mencionadas arriba no parecen participar en la modulación vocal, lo que abre la pregunta de cómo estos circuitos adquirieron una función vocal en los homínidos primitivos. Parte de la respuesta a esta cuestión ha surgido del estudio del control neuronal de los movimientos manuales, en particular de la capacidad de agarrar objetos en primates no humanos. En estos estudios se descubrieron las llamadas “neuronas espejo”, que mostraban activación no sólo cuando el animal cogía un objeto, sino también cuando veía a otro individuo realizar esta misma acción (fuera un macaco o un humano; véase Rizzolatti y Craighero, 2004). Estos circuitos fueron propuestos por algunos como la base para las capacidades de imitación y de interpretación de las acciones de otros, necesarias para el desarrollo del lenguaje. Las neuronas espejo se localizan en el lóbulo parietal inferior y en la región premotora ventral, superponiéndose en parte a la vía dorsal mencionada arriba. Por ello, se ha propuesto que los circuitos del lenguaje derivan de un circuito involucrado en el control manual, que habría sido reclutado para el control del sistema vocal (Arbib, 2005), lo cual es consistente con hipótesis previas que señalan que el lenguaje habría sido originalmente gestual, y sólo posteriormente habrían aparecido los sistemas de vocalización aprendida, dando lugar al lenguaje vocal.
Aunque coincidimos plenamente en la posibilidad del reclutamiento de circuitos temporo-parieto-frontales para el control vocal, no existe evidencia de que se haya desarrollado un lenguaje puramente manual o gestual previo al sistema vocal. La evidencia indica que, en primates y en humanos, la comunicación se basa en señales simultáneamente gestuales y vocales, que son integradas en la región de Broca. En el chimpancé, la activación del área de Broca es máxima cuando el animal efectúa gestos junto con vocalizaciones, en comparación con cualquiera de éstos por separado (Taglialatela y cols., 2011). Así, la alternativa más parsimoniosa es que, partiendo de un sistema mixto, gestual y vocal de comunicación, se fue paulatinamente robusteciendo en los homínidos primitivos un circuito sensoriomotor auditivo-vocal, que permitió la diversificación de las vocalizaciones y la transición a una comunicación predominantemente vocal. En su expansión, este circuito aprovechó la existencia de redes de control gestual y manual, que pasaron a regular procesos auditivo-vocales (Aboitiz, 2012). Creemos que esta propuesta es la que mejor sintetiza la evidencia acerca del origen neuroanatómico del habla.
Referencias
Aboitiz, F., y García, R. (1997). The evolutionary origin of the language areas in the human brain. Brain Research Reviews, 25, 381-396.
Aboitiz, F. (2012). Gestures, vocalizations and memory in language origins. Frontiers in Evolutionary Neuroscience, 4:2. doi:10.3389/fnevo.2012.00002.
Arbib, M. A. (2005). From monkey-like action recognition to human language: An evolutionary framework for neurolinguistics. Behavioral Brain Sciences, 28, 105–167.
Baddeley,A. D., Papagno, C., y Vallar, G. (1988). When long-term learning depends on short-term memory. Journal of Memory and Language, 27, 586–595.
Frey, S., Campbell, J. S. W., Pike, G. B., y Petrides, M. (2008). Dissociating the human language pathways with high angular resolution diffusion fiber tractography. Journal of Neuroscience, 28, 11435–11444.
Rilling, J. K., Glasser, M. F., Preuss, T. M., Ma, X., Zhao, T., Hu, X., y Behrens, T. E. J. (2008). The evolution of the arcuate fasciculus revealed with comparative DTI. Nature Neuroscience, 11, 426–428.
Rizzolatti, G. W., y Craighero, L. (2004). The mirror-neuron system. Annual Review Neuroscience, 27, 169-192.
Taglialatela, J. P., Russell, J. L., Schaeffer, J. A., y Hopkins, W. D. (2011), Chimpanzee vocal signaling points to a multimodal origin of human language. PLoS One, 6, e18852.
Manuscrito recibido el 28 de marzo de 2012.
Aceptado el 31 de mayo de 2012.
