
Carlos Del Río-Bermúdez (a) y Karl Ægir Karlsson (b)
(a) Universidad de Granada, España
(b) Reykjavík University, Islandia

(cc) Carlos del Río.
A pesar de los esfuerzos por comprender los mecanismos fisiológicos y funciones del sueño, el por qué pasamos más de un tercio de nuestra vida durmiendo sigue siendo uno de los grandes misterios de la neurociencia actual. Sin embargo, el uso de nuevos modelos animales simples está generando novedosos e importantes resultados sobre aspectos genéticos, fisiológicos y conductuales del sueño. El pez cebra (Danio rerio) constituye un modelo idóneo para el análisis del sueño a nivel sistémico y molecular, y sus características fisiológicas permiten la extrapolación de estos hallazgos a especies filogenéticamente superiores, como el ser humano.
Los mecanismos fisiológicos y funciones del sueño constituyen una de las principales incógnitas de la neurociencia actual. Sea como fuere, los resultados más reveladores en el área en los últimos años han estado ligados a la utilización de modelos animales de sueño, simples y bien definidos y caracterizados. Por ejemplo, el estudio del sueño en la mosca de la fruta (Drosophila melanogaster) ha permitido la identificación de una línea genética (minisleep) caracterizada por una reducción drástica del tiempo total de sueño necesario debido a una mutación en un gen que codifica un canal de potasio voltaje-dependiente (Cirelli y col., 2005). Aunque los autores no encontraron déficits asociados a la reducción de sueño en esta línea de moscas, su vida era más corta. El pez cebra (Danio rerio) constituye un organismo simple idóneo para el análisis del sueño a nivel sistémico y molecular, tanto por razones prácticas como teóricas (Hendricks, Sehgal y Pack, 2000; Zhdanova, 2011).
Entre las facilidades prácticas del trabajo con el pez cebra destacan su precio, disponibilidad y la rapidez con la que se reproducen, crían y desarrollan. La transparencia de sus tejidos en las primeras fases del desarrollo hace muy sencilla la observación y manipulación directa del sistema nervioso (véase la Figura 1). Del mismo modo, la administración de fármacos y otras sustancias no requiere del conocimiento de complejas técnicas fisiológicas, como ocurre con otros modelos (ratas y ratones), ya que únicamente basta con disolver estas sustancias en el medio natural del animal. En este sentido, destaca el potencial del pez cebra para el descubrimiento de nuevos fármacos y su impacto en el sistema nervioso central y en el sueño (para más información, véase el sitio web de 3Z).
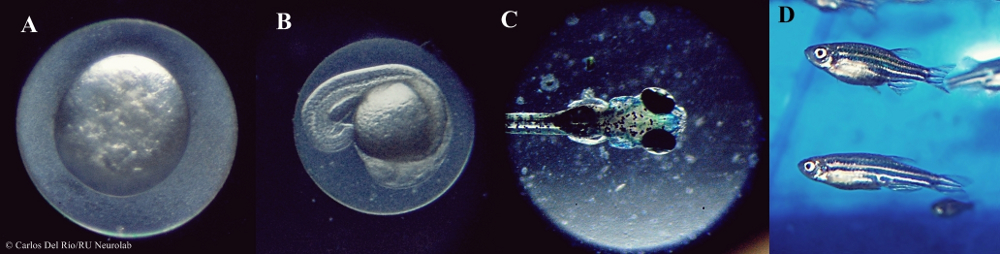
Figura 1.- Distintas fases del desarrollo del pez cebra. A: embrión en fase unicelular; B: larva, 2dpf (días post-fertilización); C: larva, 5dpf; D: pez adulto. Obsérvese la transparencia de tejidos en las primeras fases del desarrollo (A, B, C). El registro del sueño a nivel conductual se puede realizar a partir del 4-5dpf (C), cuando la vejiga natatoria está desarrollada. (cc) Carlos Del Río-Bermúdez/RU Neurolab.
La primera cuestión a responder es si los peces duermen, y si su sueño es comparable al de especies superiores. Es complicado determinar si los peces duermen atendiendo a los criterios estrictos de sueño en mamíferos (con patrones coordinados de actividad electroencefalográfica, electrooculográfica y electromiográfica). Otra postura más coherente es centrarse en parámetros conductuales del sueño más básicos, a saber: a) elevado umbral de excitabilidad; b) espontaneidad en su ocurrencia y rápida reversibilidad hacia la vigilia; c) adopción de una postura típica; y d) regulación homeostática (þorsteinsson y Karlsson, 2009). Según estos parámetros, el pez cebra presenta un estado comparable al sueño en mamíferos. Además, los neurotransmisores principales implicados en la regulación del sueño en humanos han mostrado tener un papel similar en el sueño del pez cebra (p.ej., Kaslin y Panula, 2001).
Al atender fundamentalmente a criterios conductuales del sueño (i.e., movimiento), el registro del sueño en el pez cebra a lo largo de su ciclo vital (a partir del día 4-5 post-fertilización) es relativamente simple. Para ello, se necesita únicamente una cámara de video con infrarrojos, un software de seguimiento online y un sistema de iluminación automático.
El registro conductual del sueño junto a técnicas de manipulación genética permiten estudiar directamente el papel de genes concretos en la regulación del sueño en este modelo (para consultar la secuencia genómica del pez cebra, véase The Danio rerio Sequencing Project). El uso de morfolinos (moléculas sintéticas que bloquean el proceso de traducción del ARN) permite “desactivar” selectivamente genes de interés, generando peces “knockout/knockdown” con un fenotipo conductual alterado. El morfolino es inyectado de forma masiva cuando los embriones se encuentra en fase unicelular (véase la Figura 2).
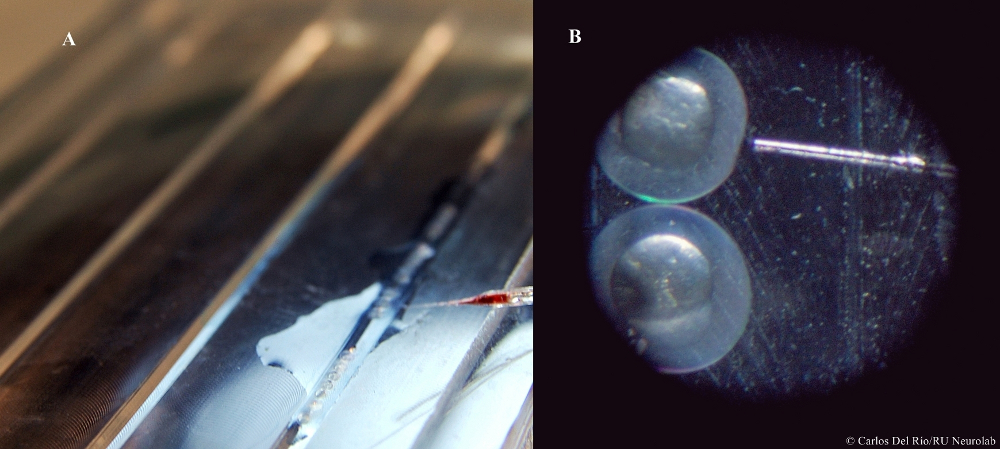
Figura 2.- Procedimiento de nanoinyección de morfolino en embriones de peces cebra. Un gran número de embriones en fase unicelular son colocados en la bandeja de inyección (A). Posteriormente, se procede a la inoculación del morfolino en la yema del embrión, bajo vista microscópica (B). (cc) Carlos Del Río-Bermúdez/RU Neurolab.
Siguiendo este paradigma hemos iniciado una línea de investigación que se centra en el efecto sobre el sueño de la desactivación temporal del gen Kcna2, que codifica una de las subunidades de un canal de potasio voltaje-dependiente (Kv 1.2), relacionada previamente en ratones con reducciones del sueño no-REM (NREM, el sueño sin movimientos oculares rápidos; Douglas y col., 2007). Los resultados preliminares señalan que estos peces knockout presentan menor tiempo total de sueño durante la noche y fenotipos asociados a la epilepsia (convulsiones), lo cual tiene sentido si consideramos que la desactivación de Kcna2 conlleva la reducción del umbral de excitabilidad neuronal.
En cuanto al papel del sueño en diversas funciones cognitivas, la investigación con el pez cebra ha generado importantes hallazgos. Por ejemplo, Rawashdeh, Hernandez de Borsetti, Roman y Cahill (2007) entrenaron a peces a asociar una parte del tanque iluminada con un entorno seguro, y otra parte del tanque en oscuridad con un entorno aversivo, donde se les aplicaban descargas eléctricas. Los peces, por naturaleza diurnos, cuando fueron entrenados y evaluados durante el día aprendieron a evitar las descargas más rápidamente (permaneciendo en la zona iluminada) y mostraron mayor retención del aprendizaje a largo plazo (24 horas) que los entrenados durante la noche. Este patrón fue constante a lo largo de diversas condiciones donde se manipulaba artificialmente el reloj circadiano de los peces. Esto llevó a pensar que la melatonina (hormona secretada fundamentalmente por la noche) podría ser responsable de estas fluctuaciones circadianas. En efecto, los autores mostraron que la posterior administración de melatonina durante el día producía resultados similares a los nocturnos, revelando el papel de esta hormona en la obstaculización de formación de memorias durante la noche.
Una de las piezas clave para resolver el enigma del sueño sería encontrar un principio (quizás a nivel molecular) que unifique su función a lo largo del desarrollo en diferentes especies (þorteinsson y Karlsson, 2009). Los vertiginosos avances en el estudio del sueño en el pez cebra nos ayudarán a comprender aspectos concretos de la fisiología del sueño en humanos, si no a desvelar el misterio del por qué dormimos.
Referencias
Cirelli, C., Bushey, D., Hill, S., Huber, R., Kreber, R., Ganetzky, B. y Tononi, G. (2005). Reduced sleep in Drosophila Shaker mutants. Nature, 434, 1087-92.
Douglas, C. L., Vyazovskiy, V., Southard, T., Chiu, S.-Y., Messing, A., Tononi, G. y Cirelli, C. (2007). Sleep in Kcna2 knockout mice. BMC Biology, 5, 42-53.
Hendricks, J. C., Sehgal, A. y Pack, A. I. (2000). The need for a simple model to understand sleep. Progress in Neurobiology, 61, 339-351.
Kaslin, J. y Panula, P. (2001). Comparative anatomy of the histaminergic and other aminergic systems in zebrafish (Danio rerio). Journal of Comparative Neurology, 440, 342–377.
Rawashdeh, O., Hernandez de Borsetti, N., Roman, G. y Cahill, G. M. (2007). Melatonin suppresses nighttime memory formation in zebrafish. Science, 318, 1144-1146.
þorsteinsson, H. y Karlsson, K. Æ. (2009). Is sleep beyond our control? The Open Sleep Journal, 2, 48-55.
Zhdanova, I. V. (2011). Sleep and its regulation in zebrafish. Reviews in the Neurosciences, 22, 27-36.
Manuscrito recibido el 8 de octubre de 2012.
Aceptado el 19 de noviembre de 2012.
