
Diana López Barroso (a,b), Antoni Rodríguez Fornells (a,b,c), Ruth de Diego Balaguer (a,b,c)
(a) Cognition and Brain Plasticity Group [Bellvitge Biomedical Research Institute]- IDIBELL, España
(b) Dept. De Psicología Básica, Universidad de Barcelona, España
(c) Institució Catalana de Recerca I Studis Avançats, España

(dp) Frances Clare.
La capacidad humana para aprender nuevas palabras es muy variable. En este trabajo mostramos que dicho aprendizaje se basa en una eficiente comunicación entre las regiones cerebrales que controlan el movimiento y las que procesan la información auditiva. Además, esta conexión anatómica, que tiene lugar a través del fascículo arqueado en el hemisferio izquierdo, no está igualmente desarrollada en todas las personas, lo que explicaría las diferencias individuales que existen a la hora de aprender las palabras de una lengua.
El aprendizaje de nuevas palabras tiene lugar a lo largo de toda la vida y, además, es un requisito necesario al aprender una segunda lengua. Esta capacidad parece ser única en los humanos, ya que la capacidad de aprendizaje de palabras en las especies más cercanas es muy limitada. Aprender palabras no sólo marca la diferencia con otras especies, sino que existe mucha variabilidad interindividual a la hora de aprender nuevas palabras. Sin embargo, existe poco conocimiento sobre las causas de esta variabilidad.
Se ha propuesto que la capacidad de aprender nuevas palabras depende de la integración que se hace, a nivel cerebral, de la representación auditiva (el “cómo” suena la palabra) y motora (los movimientos articulatorios requeridos para producir dicho sonido) (Hickok y Poeppel, 2007; Rodriguez-Fornells, Cunillera, Mestres-Missé y De Diego-Balaguer, 2009). Este proceso se conoce como integración audio-motora. Así, una vez tenemos la representación sensorial y motora de la nueva palabra, es posible asociar dicha representación multisensorial al significado. Sin embargo, las bases cerebrales precisas que permiten la integración audio-motora que sustentarían el aprendizaje de nuevas palabras no se habían establecido hasta el estudio de López-Barroso y col. (2013) que describiremos en este artículo.
En el cerebro, la interacción entre regiones distantes tiene lugar gracias a la conectividad estructural, conformada por conjuntos de axones. Las regiones cerebrales en las que hay conjuntos de axones se conocen como regiones de sustancia blanca, debido a la mielina que envuelve a los axones y que permite una eficiente transmisión del potencial de acción. Así pues, los grupos de axones hacen el papel de “carreteras” que permiten el tránsito de información de una región a otra, y la mielina que los envuelve contribuye a una mayor o menor eficacia en esa transmisión. Se ha propuesto que el principal fascículo de sustancia blanca que media el proceso de integración audio-motora, y que ha de ser crucial para el aprendizaje de palabras, es el fascículo arqueado (Schulze y col., 2012; Rodríguez-Fornells y col., 2009). El fascículo arqueado, o ruta dorsal del procesamiento del lenguaje, es un haz de sustancia blanca que, como se puede ver ilustrado en la Figura 1 (derecha), conecta regiones del lóbulo temporal (partes posteriores y mediales del giro temporal superior y medio, incluyendo el área de Wernicke) con regiones del lóbulo frontal (el giro inferior frontal o área de Broca, y la corteza premotora). Concretamente, las regiones temporales estarían relacionadas con el procesamiento auditivo, mientras que las regiones frontales sustentarían el procesamiento motor. De existir dicha relación, las diferencias individuales en la estructura de este fascículo (que afectarían a la eficiencia de la transmisión de la información de una región a otra) deben de afectar a la capacidad de aprender palabras. El fascículo arqueado conecta de forma directa las arriba mencionadas regiones fronto-temporales, pero también lo hace de forma indirecta a través de dos segmentos: uno anterior que conecta regiones frontales con la corteza parietal inferior; y uno posterior que conecta la parte parietal con la temporal (Catani, Jones y Ffytche, 2005). Además, la conectividad fronto-temporal puede tener lugar a través de un haz de sustancia blanca alternativo que transcurre por regiones más inferiores del cerebro y conecta también regiones más anteriores del lóbulo temporal: la ruta ventral, la cual ha sido relacionada previamente con procesos semánticos.
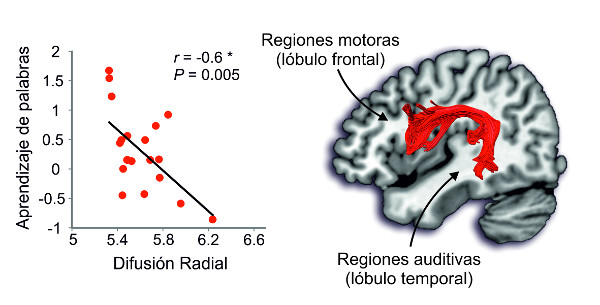
Figura 1. – Izquierda: Diagrama de correlación entre el nivel de aprendizaje de palabras (eje vertical) y el índice de Difusión Radial (RD) (eje horizontal), que refleja indirectamente la calidad de la mielina que envuelve a los axones o la propia cantidad de axones, afectando en última instancia a la eficiencia en la transmisión de la información entre regiones. Una mejor calidad o cantidad de mielina se reflejaría con un índice de RD bajo; Derecha: fascículo arqueado de un participante representativo.
Nuestro estudio (López-Barroso y col., 2013) investigó las rutas cerebrales que están en la base de la variabilidad individual observada en humanos al aprender nuevas palabras. Específicamente, el objetivo era comprobar cuál de las conexiones entre las regiones temporales y frontales se relacionaba con las diferencias individuales en el aprendizaje. Para ello, utilizamos la técnica de tractografía, que permite la reconstrucción virtual de las conexiones de sustancia blanca (Beaulieu, 2002). Esta técnica no invasiva, basada en la resonancia magnética, es sensible al movimiento de las moléculas de agua. Dado que en regiones de sustancia blanca el movimiento de las moléculas de agua está más restringido siguiendo la dirección de las conexiones o fibras, esta técnica es capaz de hacer una reconstrucción de dichas vías y extraer información sobre propiedades microstructurales que determinan la calidad de la sustancia blanca cerebral y, por lo tanto, de la eficiencia de la conectividad entre las regiones. Se realizaron, además, análisis de conectividad funcional entre las regiones de interés para complementar la información estructural aportada por la tractografía. El análisis de la conectividad funcional nos permitió extraer información sobre el nivel de sincronización en la actividad eléctrica entre regiones mientras los voluntarios estaban realizando la tarea de aprendizaje.
Un grupo de 27 participantes fueron escaneados mientras aprendían nuevas palabras. Concretamente, los participantes escuchaban un flujo de nueve palabras artificiales que se repetían varias veces. Estas palabras eran trisilábicas (p.ej., “pamedo”, “tilore”, etc.) y no tenían ningún significado asociado. Después del aprendizaje, se les administraba un test de reconocimiento en el que tenían que decidir, cada vez que escuchaban una palabra, si era una palabra que habían escuchado durante el aprendizaje o no. En el test se presentaban en igual número palabras que habían aparecido y palabras formadas con las mismas sílabas, pero en desorden. La tractografía permitió evaluar las propiedades de la mielina de las distintas conexiones que queríamos evaluar: los segmentos directos e indirectos del fascículo arqueado (ruta dorsal) y la ruta ventral, todos en ambos hemisferios.
Los resultados mostraron que las personas que tienen más mielinizada la ruta directa del fascículo arqueado en el hemisferio izquierdo fueron mejores aprendiendo las nuevas palabras. Como puede observarse en la Figura 1 (izquierda), se halló una correlación significativa entre la habilidad de aprender palabras y las propiedades de la mielina del fascículo arqueado. También encontramos una relación entre la conectividad funcional entre esas regiones y el aprendizaje de palabras. Esta relación no se observó ni para los segmentos indirectos entre dichas regiones, ni en el hemisferio derecho, ni en la ruta ventral. Este resultado muestra cómo diferencias en la anatomía cerebral reflejan diferencias en una capacidad cognitiva de gran importancia para el ser humano, como es el caso de aprender palabras auditivamente. Sin embargo, y afortunadamente, es preciso puntualizar que las diferencias anatómicas observadas pueden deberse tanto a factores genéticos como a factores ambientales (el mayor uso de una función que recae sobre ese fascículo causará cambios estructurales en el mismo; Zatorre, Fields, y Johansen-Berg, 2012). Además, el hecho de que haya una vía óptima de comunicación entre regiones (en este caso, el fascículo arqueado) no implica que esa función no pueda obtenerse a través de otra vía, aunque a un nivel menor (López-Barroso y col., 2011).
Los hallazgos de este estudio complementan modelos previos, aportando información única sobre la conectividad estructural que optimiza el aprendizaje de nuevas palabras. Además, las diferencias en la anatomía del fascículo arqueado entre humanos y primates no humanos sugiere que nuestra capacidad única para aprender lenguas se puede deber a la evolución de las conexiones fronto-temporales que, a su vez, permiten una mayor capacidad de integración audio-motora y la expansión de la memoria de trabajo verbal (manipular información verbal y mantenerla activa para ser memorizada; Schulze, Vargha-Khadem y Mishkin, 2012). Finalmente, la neuro-rehabilitación en pacientes con afasia debida a una lesión cerebral podría beneficiarse de este tipo de estudios de cara a adaptar los métodos de re-aprendizaje del lenguaje en función de la localización de la lesión.
Referencias
Beaulieu, C. (2002). The basis of anisotropic water diffusion in the nervous system – a technical review. NMR in Biomedicine, 15, 435–455.
Catani, M., Jones, D. K., y Ffytche, D. H. (2005). Perisylvian language networks of the human brain. Annals of Neurology, 57, 8–16.
Hickok, G., y Poeppel, D. (2007). The cortical organization of speech processing. Nature Reviews. Neuroscience, 8, 393–402.
López-Barroso, D., Catani, M., Ripollés, P., Dell’Acqua, F., Rodríguez-Fornells, A., y De Diego-Balaguer, R. (2013). Word learning is mediated by the left arcuate fasciculus. Proceedings of the National Academy of Sciences of the United States of America, 110, 13168–13173.
López-Barroso, D., De Diego-Balaguer, R., Cunillera, T., Camara, E., Münte, T. F., y Rodríguez-Fornells, A. (2011). Language learning under working memory constraints correlates with microstructural differences in the ventral language pathway. Cerebral Cortex, 21, 2742–2750.
Rodriguez-Fornells, A., Cunillera, T., Mestres-Missé, A., y De Diego-Balaguer, R. (2009). Neurophysiological mechanisms involved in language learning in adults. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 364, 3711–3735.
Schulze, K., Vargha-Khadem, F., y Mishkin, M. (2012). Test of a motor theory of long-term auditory memory. Proceedings of the National Academy of Sciences of the United States of America, 109, 7121–5.
Zatorre, R. J., Fields, R. D., y Johansen-Berg, H. (2012). Plasticity in gray and white: Neuroimaging changes in brain structure during learning. Nature Neuroscience, 15, 528–36.
Manuscrito recibido el 2 de octubre de 2013.
Aceptado el 5 de noviembre de 2013.
